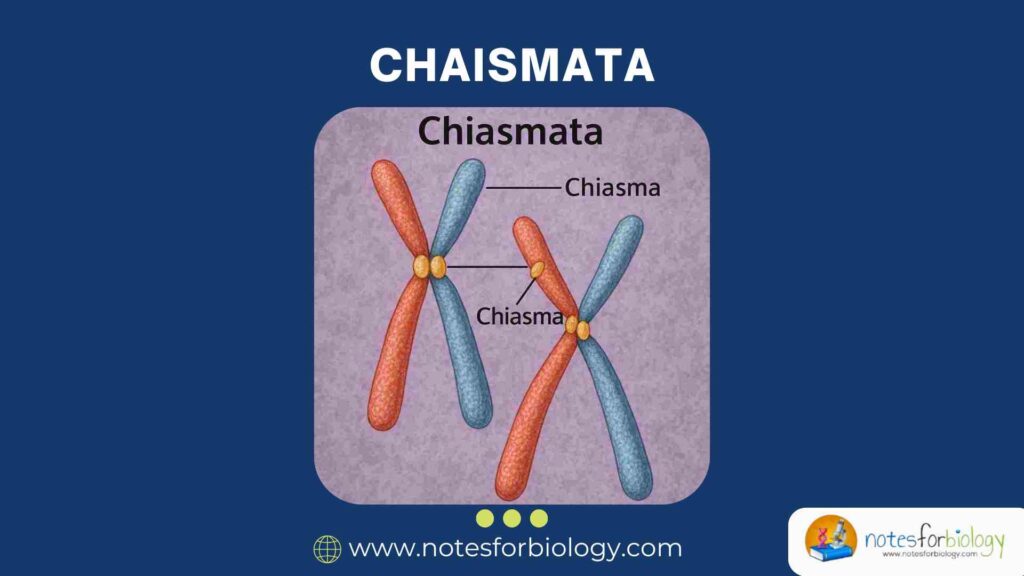
Chromosomes undergo highly regulated movements and interactions during meiosis, ensuring accurate distribution of genetic material to gametes. One of the most important features of this process is the formation of chiasmata the visible physical connections between homologous chromosomes.
This article will explore the nature, formation, and significance of chiasmata, their role in genetic recombination, and their contribution to evolutionary biology and heredity. It will also examine their involvement in genetic disorders, experimental study methods, and recent research findings.
Table of Contents
Overview of Chromosomal Behavior During Meiosis
Meiosis is a type of cell division that reduces the chromosome number by half, resulting in the formation of haploid gametes. It consists of two consecutive cell divisions: meiosis I and meiosis II.
During meiosis I, homologous chromosomes pair and exchange genetic material through a process called crossing over. Chiasmata represent the visible evidence of this genetic exchange and play a crucial role in maintaining chromosomal stability and promoting genetic variation.
Importance of Genetic Recombination in Evolution and Heredity
Genetic recombination during meiosis is essential for generating diversity within populations. The formation of chiasmata, which marks the points of genetic exchange, ensures that offspring inherit different combinations of parental traits.
This variation contributes to the evolutionary adaptability of species and is fundamental to heredity, as it shuffles alleles to create new gene combinations for each generation.
What is Chiasmata?
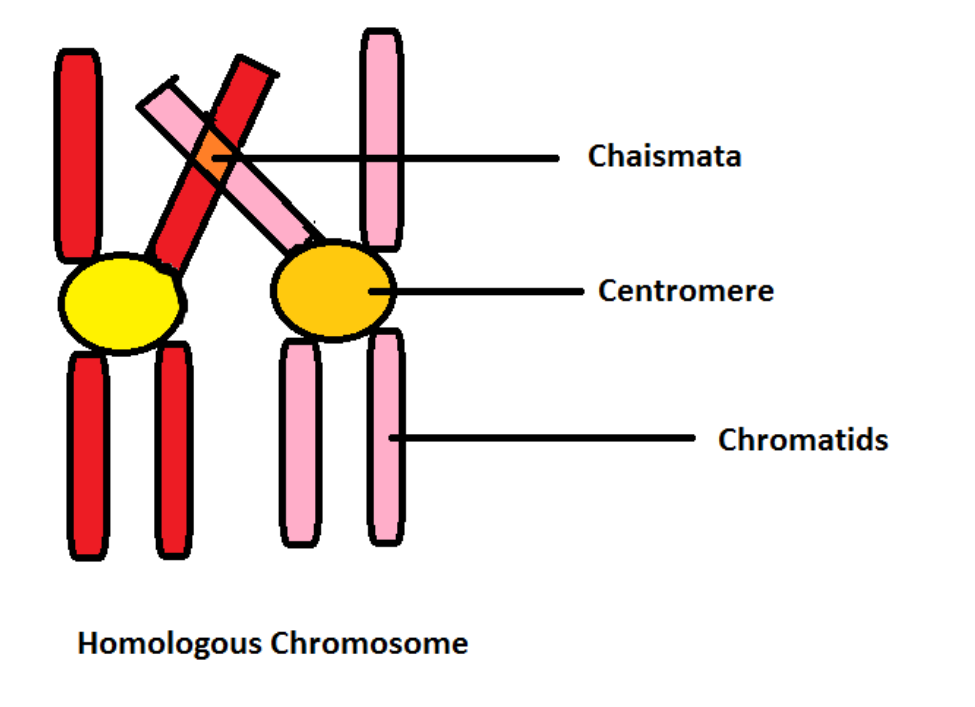
The formation of chiasmata is a defining event in meiosis, providing both a structural and genetic link between homologous chromosomes. These crossover points are vital for ensuring accurate segregation and promoting genetic diversity.
Definition of Chiasmata
A chiasma (plural: chiasmata) is the visible, X-shaped connection formed between homologous chromosomes during meiosis. It represents the site where non-sister chromatids exchange genetic segments through crossing over.
This physical link helps maintain the association between homologous chromosomes until their proper separation during anaphase I.
Discovery and Historical Background
The concept of chiasmata emerged from early microscopic observations of meiotic cells in the early 20th century. German cytologist Franz Janssens is credited with proposing the idea of chiasmata as physical crossover points in 1909.
His hypothesis, later confirmed through cytogenetic studies, provided the first evidence supporting the chromosomal theory of inheritance and the mechanism of genetic recombination.
Formation of Chiasmata
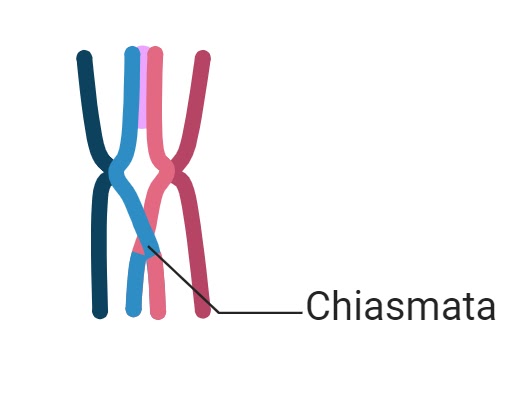
They are formed as a result of complex molecular interactions during prophase I of meiosis. The process involves the precise pairing and physical exchange of genetic material between homologous chromosomes.
Understanding how they form requires familiarity with the stages of meiosis and the molecular components involved.
Meiosis Overview: Stages Relevant to Chiasmata
Meiosis I is divided into several sub-stages: leptotene, zygotene, pachytene, diplotene, and diakinesis. Chiasmata formation occurs during the pachytene and diplotene stages.
During pachytene, homologous chromosomes are tightly aligned, and crossing over takes place. By diplotene, the points of crossover become visible as chiasmata, while other areas of the chromosomes begin to repel each other.
Process of Synapsis and Crossing Over
Synapsis is the intimate pairing of homologous chromosomes facilitated by a protein structure known as the synaptonemal complex. Once paired, non-sister chromatids break and rejoin at equivalent positions, exchanging genetic segments.
The enzymatic machinery involved includes Spo11, which induces double-strand breaks, and repair proteins like RAD51 and DMC1, which mediate strand invasion and exchange.
Formation of Chiasmata During Diplotene Phase
As meiosis progresses into the diplotene stage, the synaptonemal complex disassembles, and homologous chromosomes begin to separate. However, at the sites where crossing over occurred, they remain physically connected by chiasmata.
These connections are essential for maintaining homolog alignment until anaphase I, preventing premature separation and ensuring correct segregation.
Structure of Chiasmata
They are not random attachments but exhibit specific structural features determined by chromosomal architecture and crossover events. Their location and number directly influence the outcome of meiosis.
Physical Characteristics
Under a light microscope, chiasmata appear as cross-shaped configurations between paired chromosomes. They mark the actual physical site where genetic material has been exchanged.
Multiple chiasmata can form on a single bivalent (paired homologs), with their positions affecting the length and alignment of chromosome arms.
Chromosomal Arrangement at the Site of Chiasma
At each chiasma, the involved chromatids exchange segments and remain intertwined until separation. The chromatids rotate and arrange themselves in a specific X- or figure-eight shape, which stabilizes the homologous pair.
This arrangement facilitates the proper distribution of chromosomes to daughter cells by providing mechanical tension required for their alignment and disjunction during anaphase I.
Molecular Mechanisms Underlying Chiasma Formation
The molecular basis of chiasma formation involves coordinated action of enzymes and structural proteins. Double-strand breaks induced by Spo11 initiate the process, followed by homologous recombination mediated by proteins like RAD51, DMC1, and MSH4/MSH5.
The synaptonemal complex provides a scaffold for aligning homologs and facilitates strand exchange, while cohesion proteins like Rec8 maintain sister chromatid association until anaphase.
Functions and Significance
The presence of them is critical for both mechanical and genetic aspects of meiosis. Without them, homologous chromosomes might fail to segregate correctly, leading to severe genetic consequences.
Ensuring Accurate Chromosomal Segregation
They hold homologous chromosomes together until they are correctly oriented on the meiotic spindle. This ensures that each daughter cell receives one complete set of chromosomes, preventing aneuploidy.
Without them, homologs might segregate randomly or prematurely, resulting in gametes with abnormal chromosome numbers.
Facilitating Genetic Variation Through Crossing Over
The primary genetic significance of them lies in facilitating crossing over, which generates new combinations of alleles on chromosomes. This shuffling of genetic material contributes to genetic diversity within populations.
It also allows for the independent assortment of linked genes, promoting evolutionary adaptability and species survival.
Preventing Nondisjunction Events
They serve as mechanical anchors that resist premature separation of homologs. This tension ensures correct spindle attachment and reduces the risk of nondisjunction, where chromosomes fail to separate properly.
Such errors can lead to conditions like Down syndrome, Klinefelter syndrome, and Turner syndrome in humans.
Factors Influencing Chiasma Formation
The number, position, and frequency of chiasmata are influenced by a combination of molecular, genetic, environmental, and species-specific factors.
Role of Cohesin and Synaptonemal Complex
Cohesin proteins, particularly Rec8, maintain cohesion between sister chromatids and facilitate synapsis during meiosis. The synaptonemal complex, a tripartite protein structure, promotes close alignment of homologs and stabilizes crossover sites.
Both structures are essential for successful chiasma formation and proper chromosomal behavior.
Genetic and Environmental Influences
Genetic factors such as mutations in recombination-related genes can reduce chiasma frequency or alter crossover patterns. Environmental factors like temperature, radiation, and chemical exposure can also influence crossover rates.
For example, high temperatures have been shown to increase crossover frequency in some plant species.
Species-Specific Differences in Chiasma Frequency
Different species exhibit characteristic numbers and distributions of chiasmata. Plants typically display higher chiasma frequencies than animals, and certain chromosomes within an organism may have preferential crossover hotspots.
These differences reflect variations in genome size, chromosome structure, and evolutionary adaptations.
Types of Chiasmata
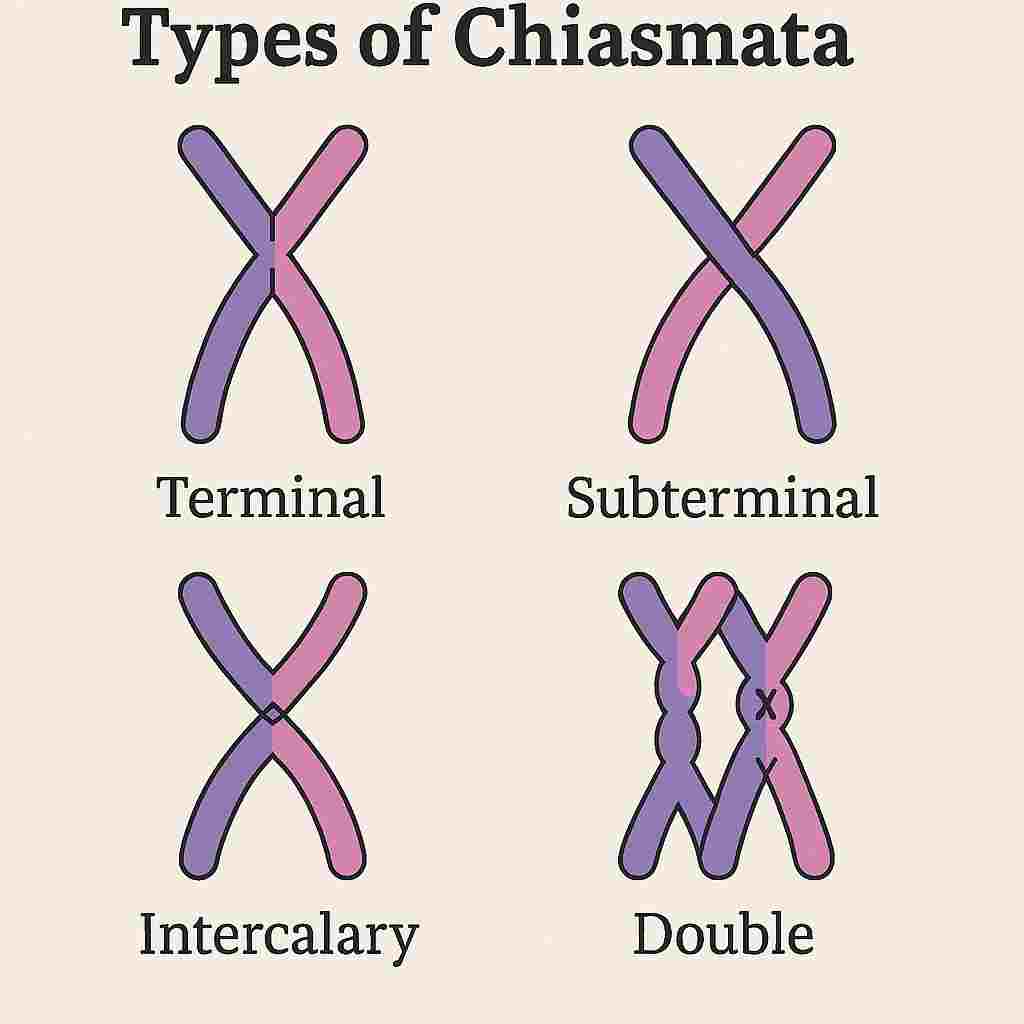
Based on their position and number on the chromosomes, chiasmata can be classified into several types. Understanding these variations aids in interpreting meiotic behavior and cytogenetic analyses.
Terminal Chiasmata
Terminal chiasmata form near the ends (telomeres) of chromosomes. They are the most common type and often occur after distal crossing over events.
These contribute to chromosome stabilization during metaphase I and ensure proper segregation.
Interstitial (Interstitial) Chiasmata
Interstitial chiasmata occur along the internal regions of chromosome arms. Although less frequent than terminal chiasmata, they are important for gene exchange within central chromosomal regions.
They help shuffle genetic material located away from telomeres and centromeres, increasing the likelihood of genetic diversity.
Multiple Chiasmata
Some bivalents may have two or more chiasmata, especially in species with large or metacentric chromosomes. Multiple chiasmata enhance genetic variation by enabling multiple crossover events along a single chromosome pair.
They also stabilize homolog alignment and reduce the risk of nondisjunction.
Chiasmata and Genetic Disorders
Errors in the formation or resolution of chiasmata can have serious genetic consequences. Faulty crossover events may lead to improper segregation of chromosomes, resulting in gametes with abnormal chromosome numbers or structures.
This section explores how chiasmata-related abnormalities can give rise to human genetic disorders and the mechanisms behind these errors.
Errors in Chiasma Formation
During meiosis, if they form incorrectly or in insufficient numbers, homologous chromosomes may fail to align or segregate properly. This can lead to nondisjunction the failure of chromosomes to separate during cell division.
Such events increase the risk of producing aneuploid gametes, which carry too many or too few chromosomes. Factors like mutations in recombination enzymes, environmental stressors, or disruptions in the synaptonemal complex may contribute to faulty chiasma formation.
Consequences in Human Genetic Diseases (e.g., Down Syndrome)
Nondisjunction caused by absent or improperly formed chiasmata is a known contributor to several chromosomal disorders. The most well-known example is Down syndrome (trisomy 21), resulting from the failure of chromosome 21 homologs to separate in meiosis I or II.
Other disorders linked to chiasmata errors include Turner syndrome (monosomy X), Klinefelter syndrome (XXY), and Patau syndrome (trisomy 13). These conditions illustrate the crucial role of chiasmata in preserving chromosomal balance in gametes.
Experimental Techniques to Study it
Cytogenetic techniques have long been used to visualize and study chiasmata in meiotic cells. Advances in microscopy and molecular biology have refined these methods, offering deeper insights into crossover mechanisms.
This section reviews traditional and modern approaches to studying chiasmata.
Cytogenetic Methods (Light Microscopy, Fluorescence Staining)
Classic cytogenetic techniques involve preparing chromosome spreads from meiotic cells and observing them under light microscopes. Staining with dyes like Giemsa or aceto-carmine reveals the physical structures of chromosomes and the presence of chiasmata.
Fluorescence staining with DNA-binding dyes like DAPI and fluorescent in situ hybridization (FISH) can enhance the visibility of chromosomes and specific DNA sequences, allowing for precise localization of chiasmata.
Modern Imaging Techniques
Recent innovations include high-resolution fluorescence microscopy, confocal microscopy, and electron microscopy, which provide detailed three-dimensional images of meiotic chromosomes.
Live-cell imaging using fluorescently tagged proteins has enabled researchers to observe the dynamics of chiasma formation in real-time. These advanced tools offer unprecedented opportunities to study the molecular behavior of chromosomes during meiosis.
Evolutionary Importance
They not only play a mechanical role in chromosome segregation but also have profound evolutionary implications. They contribute to the generation of genetic diversity and serve as markers for studying evolutionary relationships.
This section discusses how chiasmata influence evolution and genetic adaptation.
Role in Maintaining Genetic Diversity
The primary evolutionary function of it is to promote genetic diversity through crossing over. By exchanging genetic material between homologous chromosomes, chiasmata create new allele combinations in offspring.
This diversity enhances a population’s ability to adapt to changing environments, resist disease, and evolve over generations, making recombination a fundamental driver of natural selection.
Chiasmata as Evolutionary Markers
The frequency, position, and pattern of chiasmata vary across species and within populations. These variations can be used as cytogenetic markers to study evolutionary divergence and chromosomal evolution.
Comparative studies of chiasma distribution in related species help reconstruct evolutionary relationships and identify chromosomal changes associated with speciation.
Chiasmata in Plant and Animal Cytogenetics
They play distinct roles in plant and animal cytogenetics, with notable differences in their frequency, distribution, and biological significance. Studying these variations provides insights into species-specific meiotic mechanisms.
Comparative Frequency in Plant vs. Animal Cells
In general, plants exhibit a higher frequency of them compared to animals. This is especially evident in species with large or complex genomes, where multiple crossover events per chromosome pair are common.
In animals, the number of chiasmata tends to be more tightly regulated, often limited to one or two per homologous pair, reflecting differences in meiotic control mechanisms between kingdoms.
Case Studies in Model Organisms
Model organisms such as Arabidopsis thaliana, Drosophila melanogaster, and Mus musculus have been extensively studied to understand chiasma formation. Arabidopsis, for instance, displays variable chiasma frequencies influenced by environmental conditions.
In Drosophila, males lack meiotic recombination entirely, providing a unique model for studying alternative segregation mechanisms. Such case studies reveal the diversity of meiotic strategies across species.
Recent Research Advances
Modern molecular biology and genetics have expanded our understanding of the factors regulating chiasma formation. New research has uncovered additional layers of complexity in crossover control and its implications for human health and agriculture.
This section highlights recent breakthroughs in chiasma research and their applications.
Molecular Regulation of Crossing Over
Recent studies have identified specific proteins, such as the ZMM protein family (Zip1, Msh4, Msh5, and Mer3), that promote the formation of stable crossovers leading to chiasmata. These proteins work in conjunction with DNA repair factors to regulate crossover frequency and placement.
Understanding these molecular regulators has implications for fertility treatments, crop breeding, and the prevention of chromosomal abnormalities.
Implications for Genetic Engineering and Breeding
Manipulating chiasma frequency and distribution is of great interest in plant and animal breeding programs. By increasing recombination rates or targeting crossovers to specific genomic regions, scientists can accelerate the introduction of beneficial traits.
In genetic engineering, controlling crossover events may improve genome editing outcomes and minimize off-target effects by promoting precise homologous recombination.
Ethical and Clinical Considerations
The ability to manipulate meiotic processes, including chiasma formation, raises important ethical and clinical questions. While these technologies hold therapeutic potential, they also present risks and societal concerns.
This section examines the ethical dimensions of meiotic intervention and its medical applications.
Manipulation of Meiotic Processes in Labs
In laboratory settings, experimental techniques such as gene editing and chemical treatments can alter chiasma formation. While valuable for research and crop improvement, these interventions may have unintended consequences, including genome instability or ecological effects.
Ethical frameworks must guide the use of such technologies, particularly when applied to human cells or ecosystems.
Assisted Reproduction and Genetic Safety
In assisted reproductive technologies (ART), understanding them is critical for preventing chromosomal abnormalities in embryos. Techniques like preimplantation genetic diagnosis (PGD) rely on detecting recombination patterns to identify healthy embryos.
As reproductive technologies advance, ethical considerations regarding embryo selection, gene editing, and germline interventions become increasingly significant.
Conclusion
Chiasmata are indispensable structures in meiosis, serving as both mechanical links and genetic exchange points between homologous chromosomes. Their formation ensures accurate chromosomal segregation, promotes genetic variation, and safeguards against nondisjunction-related disorders.
Modern research continues to unravel the molecular mechanisms governing chiasma formation and their evolutionary implications. As genetic engineering and reproductive technologies evolve, understanding them will remain central to advancements in genetics, agriculture, medicine, and evolutionary biology.
Future studies will focus on harnessing the power of controlled recombination for crop improvement, fertility treatments, and gene therapy, while addressing the ethical and clinical challenges associated with manipulating meiotic processes.
Frequently Asked Questions (FAQ)
How do chiasmata contribute to genetic variation?
Chiasmata allow the exchange of genetic material between homologous chromosomes during meiosis, creating new combinations of alleles that increase genetic diversity in offspring.
At which stage of meiosis do chiasmata become visible?
Chiasmata become visible during the diplotene stage of prophase I in meiosis.
What proteins are essential for chiasma formation?
Key proteins include Spo11 (induces double-strand breaks), RAD51 and DMC1 (mediate strand invasion), and the ZMM protein group (like Msh4, Msh5, and Zip1) that stabilize crossover sites.
Related Contents
Chemical Reactions: Types, Balancing, Influencing Factors, and Reaction Rates